文献阅读:细胞分辨率全脑图谱的交互式框架
文献介绍

文献题目: An interactive framework for whole-brain maps at cellular resolution
研究团队: Daniel Fürth(瑞典卡罗林斯卡学院)、Konstantinos Meletis(瑞典卡罗林斯卡学院)
发表时间: 2017-12-04
发表期刊: Nature Neuroscience
影响因子: 19.9(2017年)
DOI: 10.1038/s41593-017-0027-7
摘要
为了解构大脑环路的结构和功能,有必要在全脑范围内生成神经元连接和活动的图谱。新方法现在能够以细胞和亚细胞分辨率大规模绘制小鼠大脑图谱。作者开发了一个框架,基于尺度不变的交互式小鼠大脑图谱,以细胞分辨率自动注释、分析、可视化和轻松共享全脑数据。该框架支持单个实验室和跨成像平台的连接和绘图项目,以及有关单个神经元分子身份的多重定量信息。作为概念证明,作者生成了皮质纹状体环路中五种主要神经元类型的比较连接图,以及基于活动的图来识别介导可卡因行为效应的中枢。因此,该计算框架提供了生成脑图所需的工具,集成了连接性、神经元身份和功能数据。
前言
大脑和神经环路的结构和功能图谱目前是神经科学领域的一项重大工作。大型项目已经启动,以绘制小鼠大脑的细胞类型及其活动、长程连接模式和微环路连接。绘制连接性和细胞类型的项目示例包括小鼠大脑架构项目、艾伦小鼠大脑连接图谱和小鼠连接组项目。所有此类大规模工作的核心挑战是需要开发和实施标准化系统来收集、分析、可视化和共享全脑数据。通过在全脑范围内对连接和神经元活动进行遗传标记等来剖析连接性和功能的实验方法的进步,现在可以实现环路的详细绘制。因此,除了大规模的合作项目外,越来越多的个体实验室也启动了雄心勃勃的脑图项目。然而,缺乏标准化和可访问的计算工具往往限制了大规模全脑绘图工作的能力和可行性。
在这里,作者提供一个开源软件解决方案来支持全脑图谱工作,以生成、组织和共享源自光学显微镜的全脑图谱项目。作者开发了 WholeBrain (http://wholebrainsoftware.org/),允许研究人员量化和空间绘制全脑实验的多维数据,并在单个标准化解剖参考图谱中比较不同实验的结果。作者还开发了 Openbrainmap (http://openbrainmap.org),以支持基于交互式网络框架的实验室内部和实验室之间的数据可视化和共享。整个计算框架被设计为稳健且灵活,允许其应用于各种成像系统(例如,宽场、共聚焦、光片)和标记方法(例如,荧光蛋白、免疫组织化学和原位杂交)。总之,这个计算框架提供了广泛的工具:用于在标准化大脑图谱中绘制标记神经元的强大图像处理管道、处理嵌套分层数据的贝叶斯统计包以及用于生成神经解剖数据的交互式表示的框架。该框架作为开源 R 包提供,作者通过分步视频教程制作了直观且具体的指导,允许在标准实验室环境中快速实施该系统。在这里,作者展示了如何利用 WholeBrain 和 Openbrainmap 通过整合多维解剖、分子和功能数据集来发现大脑结构-功能关系。
研究结果
1. 神经解剖学中的矢量和栅格表示
所有地图都是以视觉方式呈现物理世界的成果(Fig. 1a)。全面的神经解剖学框架必须能够处理可以描述为在大脑体积内空间定位的任何类型的数据,就像地理信息系统用于分析和表示映射到地球表面的不同类型的数据一样。
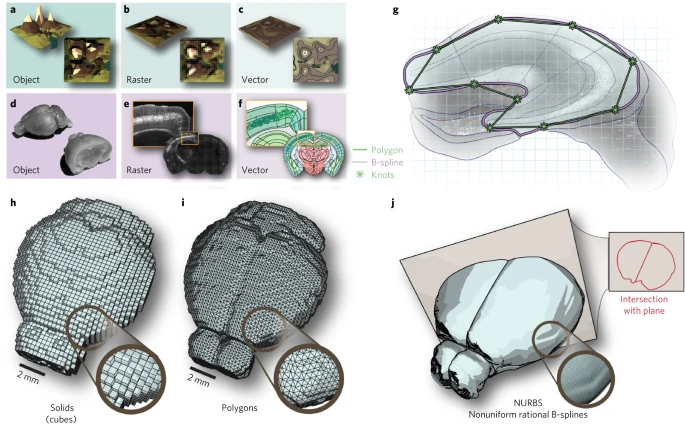
a-f. 在制图学中,对象 (a) 通常以栅格格式 (b) 或矢量格式 (c) 表示。类似地,脑组织 (d) 可以表示为栅格图像 (e) 或由矢量格式的几何点和曲线组成的图像 (f)。
g. 与从非均匀有理 B 样条曲线(NURBS,紫色)获得的区域相比,由多边形(绿色)定义的大脑区域是不可缩放的。
h–j. 在 3D 中,可以根据不同的几何基元定义参考图谱:(h) 体素可以简化为基元实体,例如立方体,(i) 多边形表面仅提供更紧凑的表面表示,并且 (j) NURBS 提供基于平滑 B 样条线的曲面表示,其中可以将曲线形式的交点计算到任意切割平面。
一般来说,地图可以使用栅格或矢量图形构建(Fig. 1b,c)。栅格图形表示网格单元结构(像素或体素)中的数据,这些数据被划分为行和列矩阵(Fig. 1b)。这种基于栅格的分析是当今神经科学的常态,例如功能性 MRI 中的全脑钙成像统计参数映射处理方法、差异基因表达(如 Allen 基因表达图谱)、以及行为条件下全脑 c-fos 表达分析(Supplementary Table 1 中可以找到最新小鼠大脑绘图方法的更多比较)。作者的目标是构建一个基于矢量图形的综合框架,以细胞分辨率表示数据,以允许通过使用点和曲线来映射特征(Fig. 1d–f)。这种方法的优点是可以在多个坐标参考系(图像像素、立体定向坐标和实际组织尺寸)中同时表示数据,而不是仅将数据转换为单个空间参考系。
2. 神经解剖学中的矢量和栅格表示
各个特征(例如神经元)的组织坐标通常使用栅格或多边形图形映射到标准化参考图谱中。相反,作者基于平滑曲线生成了成年小鼠大脑的参考图谱,这可以增强在标准化参考图谱中有效表示、共享和编辑解剖学定义的可能性。
因此,作者决定使用非均匀有理 B 样条将大脑区域表示为光滑表面(Fig. 1g)。为了生成三维 (3D) 解剖参考图谱,作者在相邻冠状切片之间放样了点。与基于实体(Fig. 1h)或多边形(Fig. 1i)的现有参考图谱相比,这种方法具有许多优点。首先,图谱是标度不变的,可以轻松应用于任何像素分辨率的图像。此功能可以比较不同显微镜系统的图像数据。其次,可以以任意角度计算图谱交集(Fig. 1j)。此功能支持在非标准角度(例如冠状或矢状)角度更好地实现区域和路径可视化的项目。第三,大脑区域的拓扑表示具有数据效率,并且项目结果可以以小格式有效共享。第四,非均匀有理 B 样条格式可以编辑大脑图谱,以适应定制和图谱改进。
该参考图谱基于 Allen 研究所小鼠参考图谱中的神经解剖学定义,从而整合了可用的神经解剖学和分子数据。例如,此功能可以从 Allen 原位杂交图谱查询任何给定立体定向坐标的基因表达。此外,参考图谱可以轻松地与其他应用程序连接,例如,通过使用定制的 3D 打印脑块进行组织切片标准化(Supplementary Fig. 1a–d),或通过生成多边形区域来自动激光捕获显微切割明确的脑核团(Supplementary Fig. 1e–j)。
3. 交互式网络界面
作者进一步开发了一个交互式网络界面,使研究人员能够以标准化格式快速可视化绘图结果,并允许他们与合作者共享数据。目的是开发一个界面,可以在标准互联网浏览器中直接执行数据可视化,而无需设置服务器。在此界面中,检测到的特征(例如细胞体以及参考图谱与组织的拟合度)可以可视化为原始栅格图像上的矢量图形叠加层(Fig. 2a–c; 请参阅 http://openbrainmap.org/example_section/ 为例),直接作为表格汇总统计进行查询(Fig. 2d,e)或在参考图谱中以立体定向坐标呈现(Fig. 2f)。
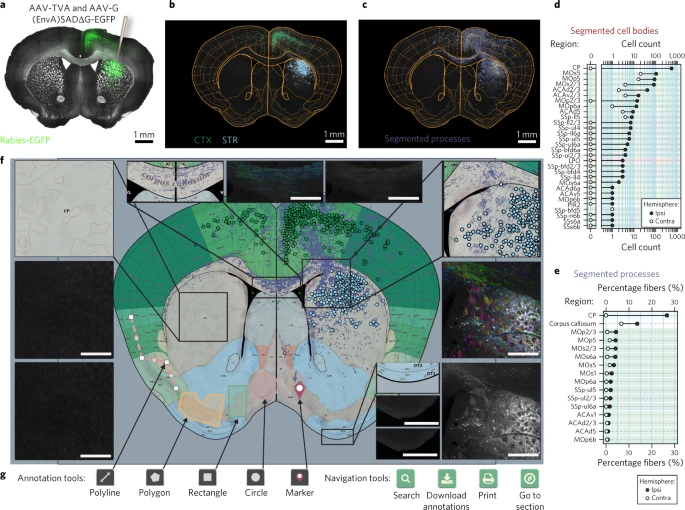
a. D1-Cre 小鼠的冠状切片,具有针对背侧纹状体的狂犬病追踪(右半球:绿色,rabies-EGFP;灰色,亮视野)。
b. 图谱区域的配准以及 1,108 个单个细胞体的分割,根据区域进行颜色编码。Striatum, STR; cortex, CTX。
c. 分割 rabies-EGFP+ 过程。
d. 分割细胞体的定量。Contra, contralateral; ipsi, ipsilateral。
e. 分割过程的定量。
f. 配准到参考图谱的结果的基于网络的矢量表示。插图:转换为参考图谱的分割结果的特写(顶部或左侧),与按色调进行颜色编码的纤维束的分割结果(中间)以及原始荧光图像(底部或右侧)进行比较。Scale bars, 500 µm。
g. 基于网络的交互式注释和导航工具。d 和 e 中缩写的大脑区域在 Supplementary Fig. 6 中定义。
为了实现快速可视化和数据共享,输出以基于浏览器的 HTML 文件形式提供,并带有关联的图像 tiles。例如,来自冠状脑切片的单个原始图像(16 bit, 300–400 Mb)的栅格数据表示为一组不同分辨率的 JPEG tiles(总共大约 20 Mb)。为了通过异步浏览器-服务器通信传输分割和配准结果,作者基于 JavaScript 对象表示法(参见 Methods)构建了空间特征及其非空间属性的开放标准格式。网络界面的设计遵循移动优先原则,支持在移动设备上使用,并为用户提供进一步绘制、编辑或定义感兴趣区域的能力(Fig. 2g)。
4. 通过多分辨率分解进行分割
分割是将标签分配给图像的不同部分以标记感兴趣的特征(例如标记的细胞体和纤维束)的过程。在大型测绘项目中,必须实施自动化的特征分割方法,该方法可以应用于所有图像,并且只需最少的手动干预。
作者使用多分辨率分解作为自动分割荧光标记的细胞体和纤维束的方法。在这种分割方法中,不同尺寸的特征(例如,细胞体和细胞突起)可以通过它们在不同空间尺度上的映射来检测。例如,顶端树突(约 1–2μm 厚)的空间尺度比细胞体(直径约 10–20μm)更小。因此,可以通过在不同尺度上指定这些特征的分割来分解树突和细胞体的遗传标记。
为了将图像分解为不同的空间尺度(Fig. 3a,b),图像数据通过由一组高通和低通滤波器组成的固定小波滤波器组发送(Supplementary Fig. 2a,b)。在每次迭代中,高通分支的输出被存储为一组称为细节系数的系数(Fig. 3c–e)。细节系数包含特定尺度的波状函数(小波)与图像中特定位置的荧光信号之间的相关性信息。通过在低通分支的输出上迭代该过程,可以获得较低分辨率级别的附加分解。每次迭代时低通分支的输出数据称为近似系数,因为在每个级别,对该分辨率级别的图像的近似变得更粗糙;细节系数表明图像中丢失了哪些细节(Fig. 3a–f)。作者通过对感兴趣的细节系数的荧光强度进行阈值化来提取各个特征(Fig. 3c–e)。然后,作者通过应用标准连通-组件算法提取每个特征,该算法按边界对连通像素进行分组(Fig. 3g,h)。
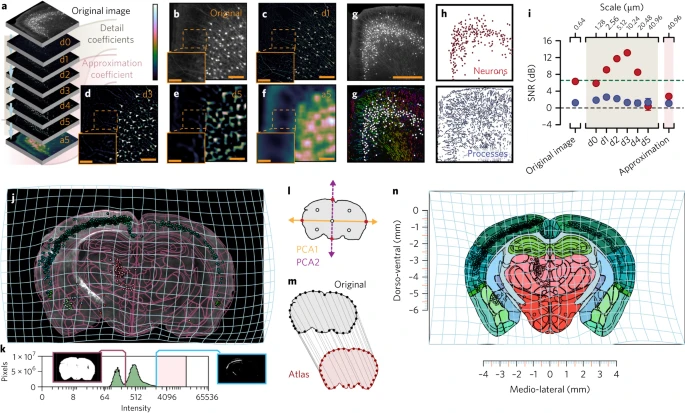
a. 小波多分辨率分解。
b. 原始图像 tile。Scale bars, 100 µm。
c. 1.28um 尺度周期的细节系数 d1。
d. 尺度周期为 10.24μm 时 d3 处的系数。
e. 尺度周期为 40.96 μm 时 d5 处的系数。
f. 近似系数。Scale bars, 100 µm。Inset scale bars, 50 µm。
g. 顶部:原始 16-bit 图像 tile。底部:分割过程,其方向按色调进行颜色编码,细胞体编码为白色圆圈。Scale bar, 500 µm。
h. 顶部:分割成细胞体的分割结果。底部:分割过程。
i. 不同细节系数的二元分割与人类注释者的比较(n = 273 个细胞体;红色,细胞体;蓝色,过程;灰色虚线,SNR 为 1:1;绿色虚线,Rose criteria SNR 5:1)。误差线:均值周围 95% 的置信区间。
j. 针对 D2-Cre 小鼠背侧纹状体的 rabies-EGFP 的图像部分(紫色线,转换后的参考图谱;浅蓝色网格,从图谱到原始图像的向后扭曲变换)。
k. 基于自发荧光的大脑切片分割(垂直紫色线)。粉色阴影区域表示可以找到细胞体的荧光强度范围。
l. 通过主成分生成对应关系(红点,与轮廓的交点;白点,中点)。
m. 原始轮廓(灰色)和参考图谱(红色)之间的 32 个对应点。
n. 分割输出的正向扭曲变换(浅蓝色网格)到立体定向空间。
为了解决该方法的灵敏性,作者在应用多分辨率分解后评估了手动定义的感兴趣区域(神经元与过程)的信噪比(SNR),并将其与原始图像中荧光信号的二值化阈值进行比较(Fig. 3g)。作者发现,神经元分割的信噪比在理论采样尺度周期为 10.24 μm 时达到峰值,大约相当于细胞体的大小(Fig. 3i)。同样,作者发现过程的 SNR 在采样周期为 2.56 μm(即顶端树突的近似直径)时达到峰值。总之,不同空间尺度的细节系数可用于识别遗传标记的细胞体和过程。
此外,多分辨率分解对于比较和整合从不同显微镜系统获得的成像结果特别有价值。本质上,可以通过分析具有相似尺度周期的尺度上的细节系数来直接比较从低分辨率和高分辨率图像中分割出的特征(Supplementary Fig. 2c–i)。因此,多分辨率分解支持跨成像平台、项目和实验室的数据收集和分割结果比较。
5. 注册到标准化图谱
图像配准是将图像组(例如,小鼠冠状脑切片)转换为另一个坐标系(例如,立体定向坐标系或参考图谱)的过程。神经影像中的传统配准算法遵循基于体素的分析。使用基于体素的图谱(例如 Allen Mouse Brain Reference Atlas 或 Waxholm space)的配准仅限于 21.5–25 μm,甚至更新的方法例如下一代通用坐标框架(CCF, v3)仍然受到限制,体素分辨率仅限于约 10 μm。为了减少计算时间,这些配准方法通常需要通过对较低分辨率的图谱进行上采样和对成像的大脑部分进行下采样来匹配图像分辨率。相比之下,作者决定通过利用大脑切片本身固有的自发荧光分割出大脑切片的轮廓来执行图像配准(Fig. 3j,k)。通过这种方式,图像配准不会引入源自与对参考图谱进行上采样相关的像素插值的人工误差。此外,无需专用一个成像通道用于配准过程,这减少了图像采集时间并有利于荧光多通道实验(多路复用)。
为了在脑组织和参考图谱之间生成界标,成像脑部分的分割轮廓被简化为一组点。为了实现这一点,首先从定义组织切片轮廓的像素位置中提取两个初始主成分(Fig. 3l)。接下来,提取主成分和组织轮廓之间的交点并将其存储为初始的第一级参考点(Fig. 3m)。默认情况下,执行四次迭代 (n = 4),从而沿着大脑部分的轮廓产生 个参考点(Supplementary Fig. 3)。两组参考点(图谱和组织切片参考点)构成一组对齐图谱和组织切片的对应点(Fig. 3m)。然后修剪这些对应点,以尽量减少组织损伤或扭曲对配准结果的影响。这些对应点用作输入,以解析方式求解薄板样条变形场的弯曲能的最小化。然后,变形场可用于生成脑组织切片(Fig. 3j)中的每个坐标与参考脑图谱(Fig. 3n)的每个坐标之间的映射。使用这种方法,可以将各个大脑部分的集合重建为成像数据的 3D 表示(Supplementary Video 1)。
当自动配准导致区域边界放置不正确时,可以手动更改或删除对应点并重新运行薄板样条算法(Supplementary Fig. 4)。为了估计测量精度如何取决于组织切片的角度和配准平面,作者比较了在冠状或矢状平面上切片的大脑的多个区域中获得的标记神经元的检测(Supplementary Fig. 5)。作为配准准确性的衡量标准,作者定义了矢状位与冠状位重建中每个解剖分区中发现的标记细胞百分比的差异。根据均方根误差的定义(Supplementary Figs. 6 and 7),所有解剖区域的平均误差为 0.29%(标准差 (s.d.) = 0.96%,n = 239 个大脑区域)。值得注意的是,重建的 3D 大脑可以以任意角度重新切片,尽管采样策略仍然会对可以获得的空间分辨率施加限制(Supplementary Fig. 8)。
6. 纤维束追踪
为了构建中尺度连接组,有必要进行纤维束追踪。纤维束是在空间中延伸(空间相干)的几何特征,在不同位置具有一组方向。作者利用相干性和方向性的特性,通过计算感兴趣的细节系数的结构张量,然后进行拓扑骨架的分割,进一步提高纤维束和细胞过程分割的信噪比(参见 Methods)。分割的拓扑骨架以及每个像素的相关方向(Fig. 4e–g)可以用作三级纤维束成像方法的输入。
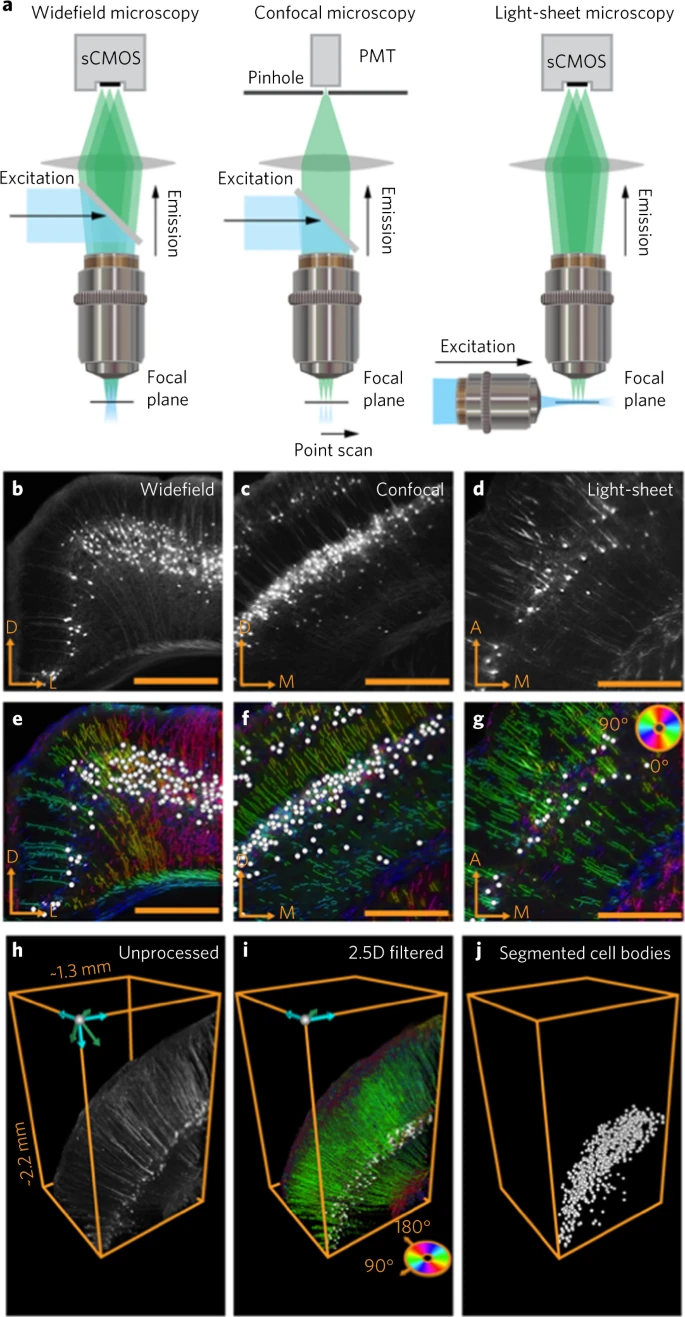
a. 框架与不同成像系统的兼容性,包括宽场、共聚焦和光片荧光显微镜。sCMOS,科学互补金属氧化物半导体相机;PMT,光电倍增管。
b-d. (b) 宽场显微镜、(c) 共焦显微镜和 (d) 光片显微镜的 rabies-EGFP 原始荧光图像。
e-g. b,c 中 (e) 宽场图像、(f) 共焦图像和 (g) 光片图像的细胞体(白色圆圈)和过程(根据角度编码)的分割结果。D, dorsal; L, lateral; M, medial; A, anterior。
h. Thy1-eYFP 小鼠中用光片显微镜成像的皮质神经元。
i. 跨 z-stacks (2.5D filter) 应用的分割算法。
j. 从归因于过程和纤维束的荧光信号中单独分割出细胞体。
确定切片组织中纤维束的身份具有挑战性。因此,需要在 3D 中进行纤维追踪,例如,在使用共焦或光片显微镜成像的完整澄清脑组织中进行纤维追踪(Fig. 4a–j)。为了处理 3D 图像堆栈(Fig. 4h),作者开发了一组 2.5D 算法,可以沿 z-stack 平面分割过程和细胞体(Fig. 4i)。这种方法在速度上与 2D 图像处理相当,但通过在每个 z 平面上组合小波和结构张量后在 z-stack 上运行附加的连通分量算法,提供了运行 3D 小波滤波器的准确性。
7. 单细胞共表达图谱
作者进一步在单神经元水平上确定荧光蛋白(例如 EGFP)成像和免疫组织化学后标记物的表达。为了绘制神经元类型的解剖分布图,作者选择使用转基因标记结合免疫组织化学来确定中间神经元的空间分布。作者使用转基因小鼠系 (Lhx6::EGFP) 结合检测表达小白蛋白 (Pvalb+) 和表达神经肽 Y (NPY+) 的中间神经元。自动图像分析结果对单个小鼠冠状脑切片中的 9,021 个标记细胞进行了分割(Fig. 5):Lhx6::EGFP:66% (n = 5,958)、PVALB::Cy5:63% (n = 5,701) 和 NPY::Cy3:15%(n = 1,342)。分割和注册到图谱的处理时间约为 30 秒,表明基于作者框架的全脑绘图项目的可扩展性。进一步证明自动化框架的优势,可以以高分辨率将这 9,021 个标记细胞体中的每一个的位置分配给 116 个独特的区域(Fig. 5a–d)。这种类型的综合单神经元映射生成详细的解剖学定义,例如,可以描述跨皮质层和不同解剖分区的神经元类型的相对分布(Fig. 5e)。作者确认了 PVALB 和 NPY (n = 0) 在 Lhx6+ 细胞中的非重叠分布(Fig. 5g,h; 37% 的皮质 NPY+ 细胞共表达 Lhx6::EGFP,n = 238,73% 的皮质 PVALB+ 细胞共表达 Lhx6::EGFP,n = 2,752)。与皮质相反,纹状体中 84% 的 NPY+ 细胞也是 Lhx6::EGFP+ (n = 101)。
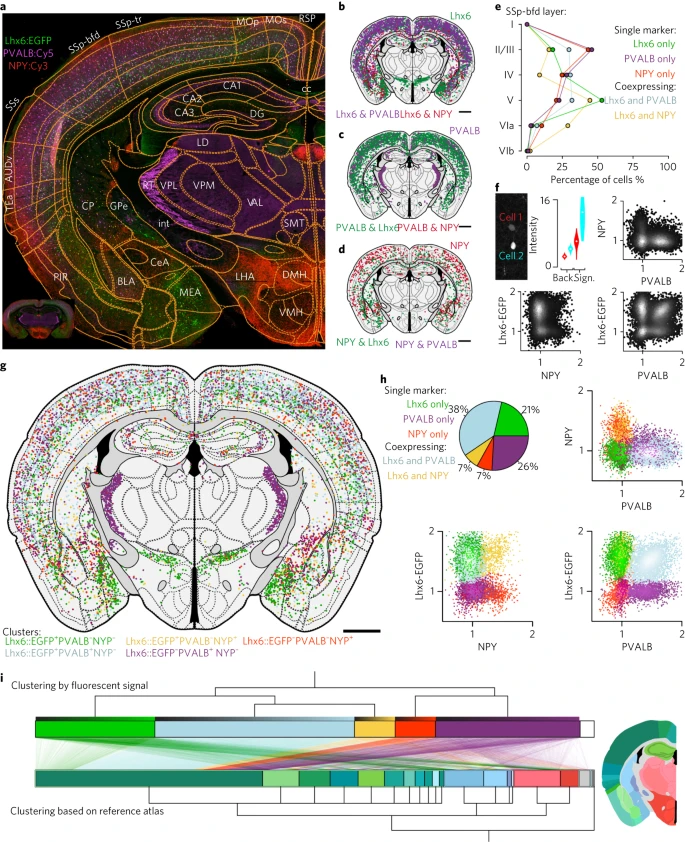
a. Lhx6::EGFP 转基因小鼠的切片(绿色),对小白蛋白(洋红色,PVALB::Cy5)和神经肽 Y (NPY::Cy3) 进行染色。Supplementary Fig. 7 中提供的缩写。
b. 分割 Lhx6::EGFP+ 细胞及其与 PVALB(紫色)或 NPY(红色)的共表达;绿色表示 Lhx6::EGFP+ 神经元对 NPY 和 PVALB 均为阴性。
c. PVALB::Cy5+ 细胞及其与 Lhx6::EGFP(绿色)或 NPY(红色)的共表达;PVALB 和 NPY 之间未发现共表达。
d. NPY::Cy3+ 细胞(红色)及其与 Lhx6::EGFP(绿色)的共表达。
e. 初级体感皮层桶状场(SSp-bfd)中细胞类型的层状分布。
f. 单细胞水平分子信号的定义(背景,n = 400 像素;信号,n = 16 像素)。
g. 将 9,021 个分割细胞体注册到参考图谱中的 116 个独特区域,并根据荧光强度分为五个离散类别。
h. 仅根据荧光强度将细胞聚类为五个离散群体(绿色,仅 Lhx6-EGFP+;洋红色,仅 PVALB+;红色,仅 NPY+;黄色,NPY+Lhx6-EGFP+;浅蓝色,PVALB+Lhx6-EGFP+)。饼图显示了重叠百分比。
i. 基于标记身份的聚类(顶部)与基于每个细胞的神经解剖位置的聚类(底部)进行比较。线条代表两种聚类类型(分子同一性与解剖学)之间映射的单个神经元。神经元根据每个 cluster 中的相对荧光强度(每个 cluster 的顶部梯度)按升序排序。解剖颜色编码显示在代表性部分,距 bregma –1.5mm(scale bar, 1 mm)。
作者进一步研究了五个不同神经元 clusters 的层状分布,发现 PVALB+(仅 PVALB)和 NPY+(仅 NPY)细胞主要定位于初级体感皮层桶状区的 II/III 层,而 Lhx6+(仅 Lhx6)和 Lhx6+NPY+ 细胞在 V 层富集(Fig. 5e)。
值得注意的是,作者的框架进一步能够量化所有标记通道的每个单细胞的平均荧光强度,从而以类似于流式细胞术中的门控的方式定量描述单细胞水平的表达水平,从而定量描述群体聚类(Fig. 5f)。使用每个荧光通道的定量信息,作者根据高斯混合模型的预期最大化将神经元聚类为五个不同的群体(Fig. 5h,i)。然后将这些 clusters 叠加在参考图谱上,以对齐图谱的分子和解剖学定义(Fig. 5h)。例如,作者发现大多数 PVALB+ 和 Lhx6::EGFP− 神经元 (PVALB+Lhx6−) 位于丘脑网状核中(Fig. 5g,i),而 Lhx6+ (Lhx6::EGFP+PVALB−NPY−) 细胞优先发现于未定带和内侧杏仁核中。这种方法同样可以通过荧光原位杂交检测单个 mRNA 分子来量化亚细胞水平上的基因表达(Supplementary Fig. 9)。
8. 使用改良狂犬病病毒和转基因小鼠追踪连接性
使用改良狂犬病病毒进行单突触连接的全脑图谱可以在中尺度水平生成细胞类型特异性连接组的大型数据集。作者应用狂犬病病毒追踪来识别皮质纹状体通路中特定神经元亚型的突触前伙伴(Supplementary Video 2)。作者使用 Cre 介导的遗传靶向方法来限制表达 Cre 的小鼠系中特定神经元类型对狂犬病病毒的摄取。作者首先使用 Camk2a-Cre 或 Gad2-Cre 小鼠分别将全脑输入映射到运动皮层 (MOp) 中的兴奋性或抑制性神经元(Fig. 6a–d)。为了靶向纹状体 (CPu) 中的主要神经元类型,作者使用 D1-Cre、D2-Cre 和 Chat-Cre 小鼠分别靶向直接通路或间接通路的中型多棘神经元以及胆碱能中间神经元(Fig. 6e–j)。作者在全脑范围内绘制了标记的突触前神经元的精确神经解剖学分布,这产生了丰富的数据集,其中包括来自 15 只小鼠和 349,959 个带注释的神经元的全脑追踪数据(Fig. 6k,l)。
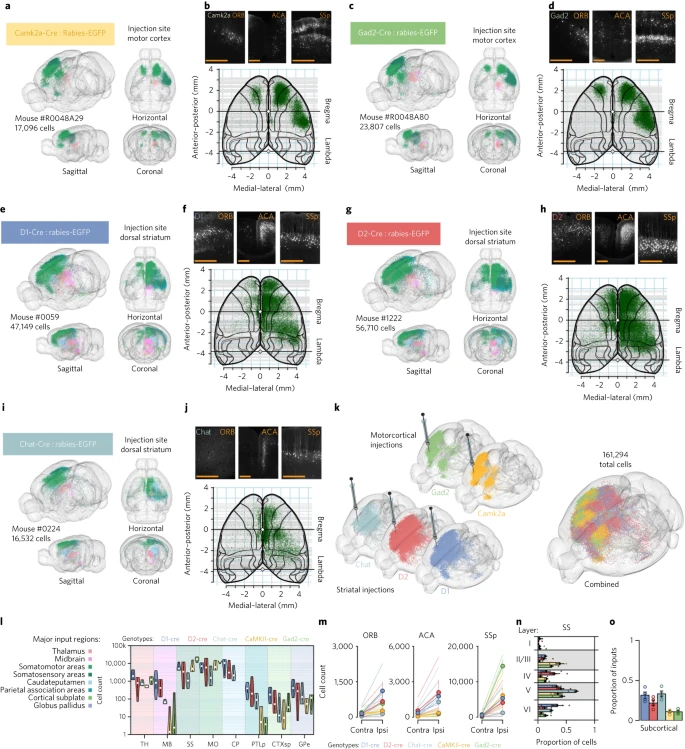
a. 通过靶向注射糖蛋白缺失的 EGFP+ EnvA 假狂犬病病毒 SADΔG-EGFP(EnvA),重建运动皮层 Camk2a 神经元的输入。EGFP+ 神经元根据解剖位置进行颜色编码。
b. 上图:ORB、前扣带区 (ACA) 和 SSp 运动皮层 Camk2a 神经元的皮层输入。底部:皮质细胞输入的皮质概览,大约位于距中线皮质表面 –2.25mm 的背腹侧上方。
c-d. 与 a 和 b 相同,但针对运动皮层中的 Gad2 神经元。
e-f. 与 a 和 b 中相同,但对于背侧纹状体中的 D1+ 中型多棘神经元。
g-h. 与 a 和 b 中相同,但对于背侧纹状体中的 D2+ 中型多棘神经元。
i-j. 与 a 和 b 中相同,但对于背侧纹状体中的胆碱能中间神经元。
k. 用于细胞类型特异性输入比较的单个大脑(来自五个不同的转基因 Cre 系)中的狂犬病 EGFP 标记的图示。大大脑是所有神经元组合在一起的复合体(5只小鼠的 n = 161,294 个神经元)。
l. 来自主要输入区域的单突触输入的小提琴图(每种基因型 n = 4 只小鼠)。白色圆圈表示中位数;粗黑线表示第一和第三四分位数;细黑线延伸 1.5 倍四分位数范围;多边形表示数据的密度估计并扩展到极值。
m. 特定区域皮质输入的偏侧性(ORB、ACA、SSp)。
n. 来自 SSp 的单突触输入的层特异性。颜色如 l 所示。
o. 来自皮层下区域的输入比例。圆圈显示单个小鼠。颜色如 l 所示。误差条: ± 1 测量标准误差。Scale bars, 500 µm。
对大量动物和实验的定量比较使我们能够寻找显示出与丘脑相似的连接模式的皮质区域(即,同时支配 MOp 和 CPu),然后可以在解剖学和功能上将其描述为上下文单元而不是作为运动皮层的输入单元。因此,作者分析了全脑追踪数据集,以专门识别表现出 MOp 和 CPu 密集靶向的皮层子区域。作者确定了三个满足上下文单元标准的候选区域:初级体感皮层(SSp)、前扣带区和眼眶皮层(ORB)。作者进一步分析了 SSp、前扣带区和纹状体 ORB 输入的细胞类型特异性组织,发现 ORB(ORBvl 和 ORBl)中的同侧输入显示出对 D1+ 纹状体细胞的优先靶向(Fig. 6m)。因此,ORB 在皮质纹状体网络中显示出独特的连接模式,优先针对纹状体 D1+ 神经元。
为了进一步研究皮质纹状体组织的层状分布,作者探索了数据集以识别所提出的细胞类型特异性连接模式。与之前依赖少量皮质神经元的研究不同,在对不同纹状体亚型进行单突触狂犬病追踪后,作者对皮质纹状体通路中的大量突触前神经元(320,055 个神经元)进行了量化。作者分析了 SSp 中细胞类型特异性皮质纹状体输入的层状分布,发现 D1+ 和 D2+ 纹状体神经元的主要输入位于 V 层,尽管作者也发现了 II/III 和 IV 层输入(Fig. 6f,h,n)。 Chat+ 纹状体神经元的输入主要位于 SSp 的 V 层和 VI 层(Fig. 6j,n)。相比之下,作者发现 MOp 中 Camk2a+ 和 Gad2+ 细胞的输入主要源自 SSp 的 II/III 层,但也可以在 V 层中找到(Fig. 6n)。
在全脑范围内比较纹状体神经元亚型的输入,作者发现与来自皮层下结构的输入相比,D2+纹状体神经元显示出来自皮层区域神经元的优先输入,而 D1+ 和 Chat+ 纹状体细胞接收更平衡的皮层-皮层下输入(Fig. 6o)。这一证据支持皮质纹状体通路以及皮质与皮质下区域的输入平衡中存在离散的细胞类型特异性连接模式。
9. 从全脑中间早期基因活动解码运动行为
为了研究通过单突触狂犬病追踪识别的皮质纹状体组织的行为相关性,作者进行了全脑功能测定。根据神经元立即早期基因(例如,c-fos)的表达来标记神经元作为神经活动的替代指标,对于识别行为过程中招募的区域和神经元类型非常有价值。
研究绘制了可卡因诱导的皮质和基底神经节环路中 c-fos 表达的图谱,最近类似的方法绘制了全脑尺度的 c-fos 表达图谱。作者选择急性可卡因给药作为一种分析方法,以绘制运动功能如何招募不同的皮质纹状体环路的图谱。作者将小鼠暴露于可卡因或盐水中,然后对 c-fos 表达进行成像,以识别表现出与运动活动增加相关的激活的神经元和区域。在开放区域中监测注射 20mg/kg 可卡因或盐水的小鼠的运动反应 20 分钟(Fig. 7a;每组 n = 4 只小鼠),并对大脑进行切片(每个大脑 M = 93.6 个切片,s.d. = 11.2 每个大脑的切片)并进行 c-fos 和 DAPI 染色(Fig. 7b–d)。
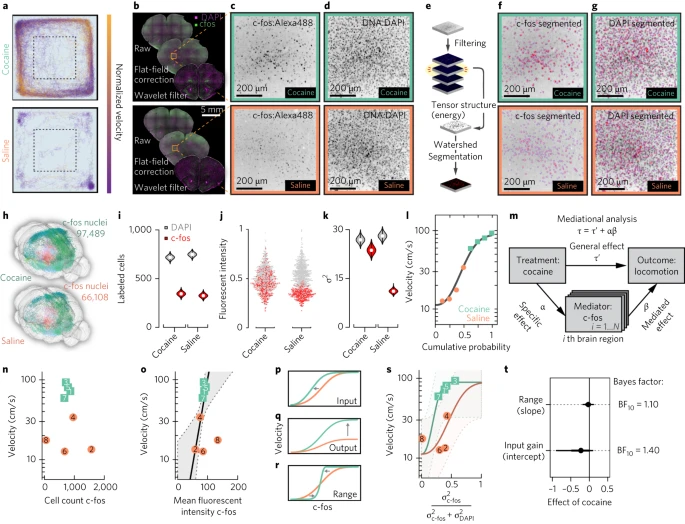
a. 对两只注射可卡因或盐水的小鼠进行旷场测试的行为轨迹追踪。
b. 来自同一小鼠的冠状切片,显示分割前预处理的主要步骤。
c. 特写 ORB 和 c-fos Alexa Fluor-488 (Alexa488) 染色。
d. 与 c 中相同,但适用于 DAPI。
e. 后过滤分割步骤包括计算张量结构和执行基于分水岭的分割。
f-g. 分割结果(红色)叠加在 (f) c-fos 和 (g) DAPI 的图像上。
h. 与 a-j 中相同的小鼠的全脑 3D 重建。
i. 从 ORB 获得的细胞计数估计值的小提琴图,如 f 和 g 所示;白色圆圈标记中线;粗黑线表示第一和第三四分位数;细黑线延伸 1.5 倍四分位距;多边形表示数据的密度估计并扩展到极值。
j. ORB 中 2,135 个单个核的标准化荧光强度。
k. ORB 中荧光强度的小提琴方差图 (σ2) 估计值;白色圆圈标记中线;粗黑线表示第一和第三四分位数;细黑线延伸 1.5 倍四分位距;多边形表示数据的密度估计并扩展到极值。
l. 8 只小鼠在旷场运动速度的累积分布(灰线,最适合四参数威布尔分布)。
m. c-fos 全脑数据的冥想回归分析。
n. 在盐水处理的(珊瑚色)和可卡因处理的(绿松石色)小鼠中,旷场测试中的速度作为眼眶皮层 c-Fos+ 核细胞计数的函数;数字表示动物识别号。
o. 速度作为平均荧光 c-fos 强度的函数;粗灰线表示当添加自发荧光作为协变量时的回归线(灰色区域,80% 可信区间)。
p-r. 通过 (p) 输入增益、(q) 输出增益或 (r) 范围压缩进行增益控制。
s. 速度作为 DAPI 标准化 c-fos 方差的函数。线条显示威布尔心理测量函数的后验估计;虚线显示 80% 的可信区间。
t. 可卡因对斜率和截距影响的后验估计。误差线,70%(粗)和80%(细)可信区间;每组 n = 4 只小鼠(可卡因、盐水)。
为了分割细胞核并确定其 c-fos 荧光强度,作者计算了后续分水岭分割的细节系数的张量能量(Fig. 7e–g and Supplementary Fig. 9)。分割算法基于空间频率分析,允许独立于信号强度进行细胞核分割,从而在宽动态范围内检测到 c-fos 信号(Fig. 7c–f)。作者在全脑尺度上绘制了 c-fos+ 神经元的位置(Fig. 7h)。
作者的分析重点是基于皮质纹状体连接模式的 ORB 活动映射。作者发现可卡因并没有显着改变 ORB 中 c-fos+ 核的数量(344 个 c-fos+ 核对 325 个, = 1,P = 0.32;Fig. 7i)。相反,作者发现可卡因显着增加了 c-fos 强度水平( = 15.21,P < 0.001;Fig. 7j)以及 c-fos 信号方差(Bartlett's K 2 = 174.21,自由度 = 2,P < 0.001;Fig. 7k)。
为了进一步绘制与可卡因给药后运动增加有功能相关的大脑区域,作者对全脑 c-fos 数据进行了线性中介分析。作者首先通过单调四参数累积威布尔分布来表征动物行为的边际分布(Fig. 7l)。作为特定大脑区域对介导效应的相对贡献的测量,作者估计了可卡因对 c-fos 表达介导的运动总效应的比例(即相对介导效应;Fig. 7m)。该分析表明,ORB 是介导行为效应的关键区域,约占相对介导效应的 7%( = 0.071,95% 置信区间:[0.00–0.898])。作者发现,c-Fos 荧光强度与速度的相关性优于绝对 c-fos+ 细胞计数(Fig. 7n,o),并且细胞计数的方差随着 c-fos+ 神经元平均数量的增加而增加(Supplementary Fig. 11)。因此,作者在后续分析中将 c-fos 荧光强度的方差标准化为 DAPI。作者询问是否可以将 ORB 中观察到的 c-fos 感应和运动之间的关系分类为一种不同的增益控制形式:输入增益、输出增益或范围压缩(Fig. 7p–r)。作者将饱和度水平固定在 90cm/s 的速度,并估计可卡因对其他两个累积威布尔参数(斜率和截距;Fig. 7s)的固定效应。使用边际累积密度函数作为先验,作者从可卡因固定效应的后验密度分布中进行采样。作者将可卡因对截距或斜率的后验固定效应与之前基于 Savage-Dickey 密度比检验的 N(0, 0.1) 的后验固定效应进行了比较。作者发现,可卡因引起的运动很可能取决于眼眶皮层 ORB 中输入增益的调制(贝叶斯因子 = 1.4),而不是范围压缩( = 1.1;Fig. 7t)。结合作者的追踪数据集(其中 ORB 被确定为优先针对 D1+ 纹状体神经元的上下文层),这些数据支持 ORB 作为介导可卡因运动效应的主要皮质纹状体中枢。
讨论
对于无脊椎动物模型生物,在开发基于电子显微镜重建的完整神经系统连接图谱和可视化方法方面取得了相当大的进展。尽管以类似规模详细重建小鼠神经系统很有价值,但获取和分析该规模的数据存在相当大的挑战。为了定义小鼠大脑中的环路组织,现在可以在细胞尺度上结合先进的转基因、标记和成像方法来生成细胞类型特异性连接的完整图谱。尽管标记方法取得了巨大进步,但用于配准和映射神经元的方法在很大程度上仍然依赖于手动检查或配准,并且无法扩展以分析大型数据集或将多个项目集成到一个数据库中。为了满足这些需求,作者开发了一个计算框架,以促进在细胞水平上对小鼠大脑环路的全面研究。
作者开发了一种计算框架,可以在全脑范围内自动分割标记神经元,而与成像方法无关。基于该计算框架,可以在尺度不变的全脑小鼠图谱中分析和呈现数据,从而允许项目和实验室之间进行快速数据比较和数据共享。该框架的一个关键优势是能够使用常用的基础设施快速可视化和共享全脑数据。为了促进该平台的集成,作者提供了常见应用程序的视频教程(http://wholebrainsoftware.org/tutorials)。
作者展示了该计算框架在狂犬病追踪数据全脑映射中的应用,这些数据源自皮质纹状体通路中不同神经元亚型,并结合使用 c-fos 映射对可卡因诱导的神经元活动的全脑定义。使用全脑狂犬病追踪,作者确定 ORB 显示皮质纹状体通路中上下文层的特征,可能塑造初级运动皮层和纹状体之间的相互作用,而且它优先针对直接通路 D1+ 纹状体神经元。作者可以进一步将这种连接模式与可卡因诱导的活动变化相结合,使用可卡因给药后单个神经元中 c-fos 水平的全脑分割,这将 ORB 确定为可卡因诱导的运动的主要调节者。
作者预计该框架的多功能性和可访问性将促进新技术和基于用户的修改的集成。例如,利用下一代测序能力的神经解剖学方法对于计算神经解剖学的发展非常重要,包括通过条形码 DNA 进行谱系追踪、通过表达 RNA 条形码对单个神经元进行连接追踪、原位测序和空间转录组学。
作者预计,对神经元、环路和活动的全脑表征的日益关注将受益于统一且直观的数据分析方法。值得注意的是,作者的框架可以整合成像领域的未来进步,纳入新的分子数据以改进神经解剖学定义并支持绘图项目中的数据共享。
注:本文为个人学习笔记,仅供大家参考学习,不得用于任何商业目的。如有侵权,请联系作者删除。

本文由 mdnice 多平台发布
相关文章:
文献阅读:细胞分辨率全脑图谱的交互式框架
文献介绍 文献题目: An interactive framework for whole-brain maps at cellular resolution 研究团队: Daniel Frth(瑞典卡罗林斯卡学院)、Konstantinos Meletis(瑞典卡罗林斯卡学院) 发表时间ÿ…...

YAML基础语言深度解析
引言 YAML(YAML Aint Markup Language,即YAML不是一种标记语言)是一种直观、易于阅读的数据序列化格式,常用于配置文件、数据交换和程序间的通信。其设计目标是易于人类阅读和编写,同时也便于机器解析和生成。在本文中…...

xcode使用
1. 界面 1.1. Build Settings,Build Phases和Build Rules三个设置项 Build Settings(编译设置): 每个选项由标题(Title)和定义(Definition)组成。这里主要定义了Xcode在编译项目时的一些具体配置 Build Phases(编译资源):用于指定编译过程中项目所链接的原文件,依赖对象,库…...

OV2640引脚的定义(OV2640 FPC模组规格书(接口线序))
OV2640是一款由Omni Vision公司生产的1/4寸CMOS UXGA(1632x1222)图像传感器。这款传感器以其小巧的体积、低工作电压和强大的功能而著称,它集成了单片UXGA摄像头和影像处理器,能够通过SCCB总线控制输出各种分辨率的8/10位影像数据…...

CTFSHOW 萌新 web10 解题思路和方法(passthru执行命令)
点击题目链接,分析页面代码。发现代码中过滤了system、exec 函数,这意味着我们不能通过system(cmd命令)、exec(cmd命令)的方式运行命令。 在命令执行中,常用的命令执行函数有: system(cmd_code);exec(cmd_…...

深入Java数据库连接和JDBC
引言 Java数据库连接(JDBC)是Java语言中用于执行SQL语句的标准API。通过JDBC,开发者可以方便地与关系型数据库进行交互。然而,直接使用JDBC API面临着数据库连接管理复杂、性能瓶颈等问题。数据库连接池作为一种解决方案,可以有效地管理数据库连接,提高应用程序的性能。…...
与长短期记忆网络(LSTM)结合的预测模型(GWO-LSTM)及其Python和MATLAB实现)
灰狼优化算法(GWO)与长短期记忆网络(LSTM)结合的预测模型(GWO-LSTM)及其Python和MATLAB实现
#### 一、背景 在现代数据科学和人工智能领域,预测模型的准确性和效率是研究者和工程师不断追求的目标,尤其是在时间序列预测、金融市场分析、气象预测等领域。长短期记忆(LSTM)网络是一种解决传统递归神经网络(RNN&a…...

电路板热仿真覆铜率,功率,结温,热阻率信息计算获取方法总结
🏡《电子元器件学习目录》 目录 1,概述2,覆铜率3,功率4,器件尺寸5,结温6,热阻1,概述 电路板热仿真操作是一个复杂且细致的过程,旨在评估和优化电路板内部的热分布及温度变化,以确保电子元件的可靠性和性能。本文简述在进行电路板的热仿真时,元器件热信息的计算方法…...

C#中多线程编程中的同步、异步、串行、并行及并发及死锁
在C#中,多线程编程是一个强大的功能,它允许程序同时执行多个任务。然而,这也带来了复杂性,特别是在处理同步、异步、串行、并行、并发以及死锁等问题时。下面我将详细解释这些概念,并给出一些C#中的示例和注意事项。 …...

【Lampiao靶场渗透】
文章目录 一、IP地址获取 二、信息收集 三、破解SSH密码 四、漏洞利用 五、提权 一、IP地址获取 netdiscover -i eth0 Arp-scan -l Nmap -sP 192.168.78.0/24 靶机地址:192.168.78.177 Kali地址:192.168.78.128 二、信息收集 nmap -sV -p- 192.…...

使用WebSocket实现log日志流的实时展示-从轮询到通知
场景介绍 最近开发一个系统,其中一个模块需要展示实时的执行过程,过程日志可能比较多。以前的方案都是前端定时轮询,比如每秒查一次后端接口,将拉取回来的日志重新展示。轮询方案简单容易实现,但是比较消耗资源&#…...
UE5 从零开始制作跟随的大鹅
文章目录 二、绑定骨骼三、创建 ControlRig四、创建动画五、创建动画蓝图六、自动寻路七、生成 goose八、碰撞 和 Physics Asset缺点 # 一、下载模型 首先我们需要下载一个静态网格体,这里我们可以从 Sketchfab 中下载:Goose Low Poly - Download Free …...

O’Reilly
--江上往来人,但爱鲈鱼美。 --君看一叶舟,出没风波里。 OReilly OReilly出版社出版的技术类图书 俗称动物系列 应该是每个技术人员的必备手册。 OReilly动物系列(中译本) 简介" 动物系列作为 OReilly 书籍的典型代表被普遍…...

优盘驱动器未格式化:数据拯救行动指南
优盘困境:驱动器未格式化的挑战 在日常的数据存储与传输中,优盘以其便携性和高容量成为了我们不可或缺的伙伴。然而,当您尝试访问优盘时,突然弹出的“驱动器未被格式化”提示却如同晴天霹雳,让人措手不及。这一状况不…...

4.Handler mappings
处理程序映射 简介 在早期版本的 Spring 中,用户需要在 Web 应用程序上下文中定义一个或多个 HandlerMapping bean 以将传入的 Web 请求映射到适当的处理程序。随着注解控制器的引入,通常不再需要这样做,因为 RequestMappingHandlerMapping…...
《学会 SpringMVC 系列 · 消息转换器 MessageConverters》
📢 大家好,我是 【战神刘玉栋】,有10多年的研发经验,致力于前后端技术栈的知识沉淀和传播。 💗 🌻 CSDN入驻不久,希望大家多多支持,后续会继续提升文章质量,绝不滥竽充数…...

深度学习项目 -7-使用 Python 的手写数字识别
一、前言 该文章仅作为个人学习使用 二、正文 项目源代码:深度学习项目 - 使用 Python 进行手写数字识别 - DataFlair (data-flair.training) 数据集:https://drive.google.com/open?id1hJiOlxctFH3uL2yTqXU_1f6c0zLr8V_K Python 深…...

MySQL —— 库,数据类型 与 表
库与基础操作 1.1 查看数据库 使用 show databases; 可以查看当前 MySQL 目前有多少个数据库 5 rows 表示有 5 行,这里是表示的是有效的数据,不包括 第一行的指引 set 表示结果集合 0.01 sec 表示这个 sql 语句一共运行了0.01 秒,一般情况…...

Java重修笔记 第二十七天 匿名内部类
匿名内部类 1. 定义:无类名(底层自动分配类名“外部类名$1”),既是类也是对象,定义在外部类的局部位置,例如方法体和代码块中,通过new类或接口并在大括号里重写方法来实现。 2. 使用场景&…...

Nero Lens 智图 - 适用于 iOS 和 iPadOS 的专业图片处理 App
首先是手机端的无损放大 App:Nero Lens 智图,适用于 iOS 和 iPadOS,不仅可以放大,还有多种 AI 图片增强功能。 使用这款 App 可以通过 AI 模型智能放大可达 400%,还有老照片去划痕、上色,抠图移除背景、照…...

用Wireshark抓包学LTE:手把手解析开机附着流程中的NAS/RRC消息
用Wireshark抓包学LTE:手把手解析开机附着流程中的NAS/RRC消息 1. LTE信令分析实战环境搭建 工欲善其事,必先利其器。在开始解析LTE信令前,我们需要搭建专业的分析环境。不同于传统教材的理论讲解,我们将从工程师视角构建完整的分…...

如何用Penpot构建完整的用户体验地图和用户旅程:7步打造完美设计流程
如何用Penpot构建完整的用户体验地图和用户旅程:7步打造完美设计流程 【免费下载链接】penpot Penpot - The Open-Source design & prototyping platform 项目地址: https://gitcode.com/GitHub_Trending/pe/penpot Penpot作为开源的设计与原型平台&…...

Llama-3.2V-11B-cot代码实例:Streamlit中图片上传与缓存机制
Llama-3.2V-11B-cot代码实例:Streamlit中图片上传与缓存机制 1. 项目概述 Llama-3.2V-11B-cot是基于Meta Llama-3.2V-11B-cot多模态大模型开发的高性能视觉推理工具,专为双卡4090环境优化。该工具通过Streamlit构建了直观易用的交互界面,特…...

从轨迹到网络:广州休闲步行空间格局刻画 | 论文全解析与方法论深度拆解
从轨迹到网络:广州休闲步行空间格局刻画 | 论文全解析与方法论拆解 原文:From trajectories to network: Delineating the spatial pattern of recreational walking in Guangzhou》 一、论文核心概览:摘要与关键词 1.1 核心摘要解析 本文的核心内容可拆解为5个核心模块,…...

OpenClaw技能分享:GLM-4.7-Flash驱动的邮件自动处理系统
OpenClaw技能分享:GLM-4.7-Flash驱动的邮件自动处理系统 1. 为什么需要自动化邮件处理 每天早晨打开邮箱,看到堆积如山的未读邮件总让人头皮发麻。作为一个小团队的负责人,我经常需要处理客户咨询、内部沟通、会议邀请等各种类型的邮件。最…...
)
百度快速排名优化技术(百度seo排名优化)
百度快速排名优化技术是一种针对搜索引擎结果页面(SERP)排名优化的技术手段,通过优化网站的内容、结构和用户体验等方面,提高网站在搜索引擎中的排名,从而获得更多的流量和潜在客户。下面,我将介绍百度快速…...

OpenClaw密码管理:nanobot安全存储与自动填充方案
OpenClaw密码管理:nanobot安全存储与自动填充方案 1. 为什么需要本地化的密码管理方案 去年的一次数据泄露事件让我彻底放弃了所有云端密码管理器。当时我使用的某知名商业工具突然弹出安全警报,提示"您的部分密码可能已被未授权访问"。虽然…...
 飞控IMU方向调整)
手搓STM32H743开源飞控系列教程---(五) 飞控IMU方向调整
1. 为什么需要调整飞控IMU方向 第一次玩飞控的朋友可能会遇到一个奇怪现象:明明把飞控板水平放在桌面上,地面站显示的姿态却歪了30度。这种情况十有八九是IMU安装方向与飞控默认设定不匹配导致的。我刚开始玩穿越机时就踩过这个坑,当时把飞控…...

OpenRocket:模型火箭仿真的全流程技术解决方案
OpenRocket:模型火箭仿真的全流程技术解决方案 【免费下载链接】openrocket Model-rocketry aerodynamics and trajectory simulation software 项目地址: https://gitcode.com/GitHub_Trending/op/openrocket OpenRocket作为一款开源的模型火箭仿真软件&…...
)
从零开始:用QGIS和PostgreSQL构建交通路线空间数据库(含Python脚本自动化技巧)
从零开始:用QGIS和PostgreSQL构建交通路线空间数据库(含Python脚本自动化技巧) 在交通规划与智慧城市建设的浪潮中,空间数据的高效管理成为技术团队的核心挑战。传统文件存储方式难以应对大规模交通网络数据的实时查询与分析需求&…...