论文阅读:《Whole-animal connectomes of both Caenorhabditis elegans sexes》
一 论文整体概述
论文下载链接:《Whole-animal connectomes of both Caenorhabditis elegans sexes》
补充信息和额外数据:https://www.nature.com/articles/s41586-019-1352-7
1. 作者期刊背景
该论文由Scott W. Emmons,David H. Hall等人著作,并发表在《Nature》杂志上。Scott W. Emmons和David H. Hall是该领域的知名专家,长期致力于线虫 C. elegans 的神经生物学研究。Emmons 实验室专注于神经回路功能,而 Hall 实验室则在电镜重建方面有卓越的贡献。这篇论文发表在《Nature》杂志上,这是一个顶级科学期刊,以发表原创、重要且具有广泛影响力的研究而闻名。发表在《Nature》上的论文通常都经过严格的同行评审,质量很高。
2. 论文大概内容
这篇论文的核心内容是:首次完整地绘制出了雄性和雌雄同体秀丽隐杆线虫 (C. elegans) 的全动物连接组(connectome) (连接组是指生物体内所有神经元及其连接的完整图谱)。
秀丽隐杆线虫(Caenorhabditis elegans,C. elegans)是一种非寄生性线虫,以大肠杆菌等微生物为食,身体透明,长度约1毫米,主要分布在温带地区的土壤中。秀丽隐杆线虫有雄性(Male)和雌雄同体(Hermaphrodite)两种性别,在自然条件下,雌雄同体占大多数,可自体受精,也可接受雄虫的精子产生后代。
秀丽隐杆线虫是一种模式生物,其神经系统相对简单,但功能多样。此前的研究已经建立了雌雄同体线虫的连接组,但雄性线虫的连接组尚未完整绘制。这两种性别线虫在神经系统结构上有一些差异,这些差异可能导致行为和生理上的显著区分,值得探讨与研究。
- 研究方法:
- 该研究利用先进的连续切片电子显微镜(serial-section electron microscopy, sSEM)技术,对雄性和雌雄同体线虫的整个身体进行了高分辨率扫描。
- 通过对大量电镜图像的人工注释和三维重建,研究人员精确地追踪了每个神经元的胞体、轴突和树突,并识别和记录了神经元之间的突触连接。
- 他们还开发了新的算法和分析工具,以处理如此庞大的数据量。
- 主要发现:
- 完整的雄性连接组: 这是该研究最主要的一个贡献,他们首次完整地绘制了雄性线虫的连接组,包括所有的神经元和突触连接。
- 不同连接组的比较: 研究发现,雄性和雌雄同体线虫的神经回路有许多相似之处,但也有显著的性别特异性差异,尤其是在与交配相关的神经回路中。
- 新发现神经元信息: 在雄性线虫中,研究人员发现了以前未知的神经元类型和连接模式。
- 详细突触连接类型: 该研究提供了有关突触连接类型的详细信息,包括化学突触和缝隙连接(即所谓的电突触)。
- 性别差异结构基础: 通过比较两种性别连接组,研究人员揭示了性别差异的神经回路结构基础,这有助于理解性别行为的神经机制。
二 论文核心内容
1. Abstract
这项研究首次完整地绘制了秀丽隐杆线虫(C. elegans)的雄性和雌雄同体的全动物连接组,为我们提供了该模式生物两种性别所有神经元及突触连接的详细图谱。研究人员利用连续切片电子显微镜技术对雄性和雌雄同体线虫进行了高分辨率扫描,并结合人工注释和三维重建技术,精确地追踪了神经元的结构和连接。
通过比较不同性别连接组,研究揭示了两者神经回路的相似之处和性别特异性差异,尤其在与交配相关的神经回路中发现了显著的差异。此外,研究还在雄性线虫中发现了先前未知的神经元类型和连接模式。这些发现不仅提供了关于性别差异神经回路结构基础的新见解,也为计算神经科学提供了宝贵的资源,促进了对神经系统功能和性别的神经机制的理解。这项工作标志着神经科学研究的一个重要里程碑,并为未来的神经回路研究奠定了基础。
动物能够进行各种各样的行为,这些行为必须被集体控制并紧密整合,以促进生存和繁殖。为了理解动物行为,不仅需要识别特定反应和动作的细胞基础和回路,还需要描述这些回路如何整合以产生有凝聚力和优先级的适应性输出。一个涵盖整个神经系统的连接图对于研究如何实现这种整合是必要的。迄今为止,已发表的源自电子显微照片的突触水平的神经图谱包括线虫秀丽隐杆线虫和太平洋索线虫、小鼠的视网膜和部分新皮层、果蝇的视觉系统、蘑菇体、运动和幼虫逃跑反应回路、环节动物扁头虫的幼虫视觉系统以及海鞘尾索幼虫的中枢神经系统。在这里,我们提出了包含线虫秀丽隐杆线虫成年两种性别神经系统的整个动物连接组。
2. Whole-animal connectomes
先前的研究已经描述了秀丽隐杆线虫神经系统主要神经中枢的连接性,例如成年雌雄同体的头部和尾部区域以及咽部,还有成年雄性的尾部区域。这些数据不仅构成了秀丽隐杆线虫神经科学研究的基础,也构成了对网络结构及其潜在动力学和控制特性的研究基础。
在这里,我们通过新的电子显微镜(EM)系列数据,增加了对雄性头部神经环和后泡神经节的神经回路重建,并重新注释了先前生成的雌雄同体的图谱。这使我们能够以多种方式扩展之前的工作(Extended Data Fig. 1, Methods and Supplementary Information 1)。这些重建是定量的,基于突触的大小(Methods, Extended Data Fig. 2 and Supplementary Information 2, 3),从而使图和网络分析成为可能。我们对数字化图像的注释方法使我们能够比之前的研究记录到更多的突触(Extended Data Fig. 1 and Methods)。
由于没有任何一个电子显微镜系列能够覆盖整个单一动物,为了生成全动物连接组,我们将来自不同重建系列的数据进行合并,并通过推断已知连接在重复区域之间的关系来填补剩余的空白。
雌雄同体连接组的图有 460 个节点(302 个神经元,132 块肌肉和 26 个非肌肉末端器官),而雄性连接组的图有 579 个节点(385 个神经元,155 块肌肉和 39 个非肌肉末端器官)。完整的细胞列表在Supplementary Information 4中给出。相应的图在雌雄同体中有 4,887 条化学(或定向)边和 1,447 条缝隙连接(或无向)边,在雄性中有 5,315 条化学边和 1,755 条缝隙连接边(Extended Data Fig. 3 and Supplementary Information 5)。它们是稀疏图,分别占所有可能边的 3.2%(对于雌雄同体)和 2.4%(对于雄性)。虽然稀疏,但如果所有边的方向都被忽略,无论是化学突触连接还是缝隙连接,这两种性别的图都是连通的,这意味着存在一条路径连接每一对节点(弱连通)。由缝隙连接引起的物理连接的比例在神经元类别中差异很大,对于非咽部中间神经元,其范围从超过 90% 到小于 5% 不等(Extended Data Fig. 3d)。
基于计算排列的连接性图的二维布局揭示了感觉信息流的路径,感觉神经元和末端器官之间少量的(1-5个)突触步骤,以及网络的前馈性质(Fig. 1; A3 and interactive versions of Fig. 1 can be found in the Supplementary Information)。节点在布局上与蠕虫神经解剖结构的显著相似性反映了经济的布线,这是包括秀丽隐杆线虫在内的神经系统常见的属性。
尽管两种性别的图布局表面上看起来不同,但仔细检查表明,对于两种性别都存在的神经元,包括头部中为头部运动和在物理及化学环境中导航提供感觉输入和决策的神经元,连接路径相似但不完全相同(see ‘Comparison of the sexes’)。在动物体内,神经元和神经突的排列方式类似(Extended Data Fig. 4)。主要的性别差异在于生殖功能,雌雄同体的外阴和子宫肌肉以及控制它们的运动神经元;而在雄性中,则是在尾部存在大量额外的神经元、性肌肉和连接,这些形成了交配的神经回路。
3. The architecture of information flow
我们使用化学突触的极性和物理连接网络的结构,利用一种能够检测网络中层次结构的算法来对性别共享的神经元和末端器官类别进行排序 (Fig. 2)。(Lists of the neuron and end-organ classes are provided in Supplementary Information 6; adjacency matrices by cell class are included in Supplementary Information 7.)
中间神经元可以大致分为四个层次,其中前三个层次和最后一个层次的划分依据不同。前三个层次的中间神经元大致呈现典型的前馈神经网络结构,所以根据某个神经元到运动神经元的距离(以突触数量衡量)和到后续层级的影响力(比如连接强度等)划分了三个层级。第四类中间神经元,即跨所有层次相互作用的中间神经元,不能适应这种分层结构。
化学连接和缝隙连接分布在整个网络中(Extended Data Fig. 6)。
两种性别共有的83个感觉神经元可以根据刺激类型、连接性和引发行为反应的性质分为六类。这些类别的输出在网络中差异性地分布(Fig. 3)。三个中间神经元层之间的化学连接形成了一个前馈环路:第三层主要靶向第二层和第一层,而第二层则靶向第一层。正如先前所指出的,前馈环路是秀丽隐杆线虫连接组中普遍存在的模体(Extended Data Fig. 7)。
由于秀丽隐杆线虫的感觉神经元通常对多种刺激做出反应,感觉整合从感觉神经元本身内部开始。网络中单个节点处汇聚和发散的程度由一个称为节点度的量来指定,它指的是附着边的数量(邻居的数量)(Extended Data Fig. 3)。考虑到细胞类别之间连接的图,大约 19 的中值度值说明了广泛的交叉连接,因此表明了整个神经系统中的汇聚和发散。发散的连接使得来自单个感觉神经元的信息有可能在两个突触步骤内到达网络中 70% 到 98% 的其他细胞。
除了到肌肉的输出外,神经系统还与非肌肉末端器官有相当多的连接,包括皮下组织、肠道、排泄系统,以及在雄性中,性腺(Extended Data Fig. 8 and Supplementary Information 5)。这些连接表明,神经系统在影响生理方面的作用很重要,不仅限于分泌突触外信号。
4. Generation of body movements
秀丽隐杆线虫在觅食和运动过程中的姿势运动是由一组 95 块体壁肌肉产生的,这些肌肉排列成四个纵向行,其中两个是背侧的,另外两个是腹侧的。在每个象限内,相邻的肌肉细胞通过缝隙连接进行电耦合(Fig. 1)。在 46 个类别中,有 154 个神经元与这些躯体肌肉有神经肌肉接头(Extended Data Fig. 9a)。在这些神经元中,我们将 17 个类别中的 108 个神经元(占体壁肌肉输入的 91%)归类为运动神经元,因为它们的大部分输出是到肌肉,而且肌肉控制似乎是它们的主要功能(Extended Data Fig. 9b)。我们对一些运动神经元的分类与之前的分类有所不同(Supplementary Information 1)。
108个运动神经元可以细分为三组。五类头部运动神经元支配身体前部区域的肌肉(URA、RME、RMD、RIV和RMH),七类腹索运动神经元支配身体其余部分的肌肉(VA、DA、VB、DB、VD、DD和AS),五类侧下方运动神经元(SAB、SMD、SMB、SIB和SIA)的轴突在靠近四个肌肉象限的侧下方束中纵向延伸,从而输出到肌肉(Extended Data Fig. 10a)。侧下方运动神经元在此之前尚未被完全描述,因为它们的轴突穿过尚未在高放大倍率下通过电镜完整成像的身体区域。对合成和释放乙酰胆碱的机制进行免疫荧光染色显示,这种神经递质沿着侧下方束广泛释放,至少在身体前三分之二的部分是这样(Extended Data Fig. 10b)。对可用显微照片的有限抽样检查证实,释放位点对应于位于含有突触前致密区的肿胀处的、向皮下组织和肌肉的双突触(Extended Data Fig. 10c, d)。功能研究表明,SMD编码一种被称为Ω形转弯的深腹侧弯曲行为的幅度;SMB影响正弦运动的幅度;SAB被认为在本体感受中起作用;而SIA神经元则参与滞眠期间的所谓“翻转行为”。
可以预期的是,蠕虫的姿势将由这三组运动神经元的总输入所产生的肌肉张力所形成。事实上,通过对四个“特征蠕虫”的形状进行求和,可以很好地描述蠕虫的姿势,这些形状分别对应于这三组运动神经元:前两个是正弦波形,对应于腹索运动神经元的前向和后向类别;第三个是全身弯曲,对应于侧下方运动神经元的潜在影响;第四个包括头部弯曲,对应于头部运动神经元。
这三类运动神经元成员的支配是复杂的。总的来说,来自 82 个类别中的 60 个类别(73%)的中间神经元和感觉神经元(126 个神经元)对运动系统(运动神经元和肌肉)有显著强度(超过三个连续的电镜切片)的突触输入(Fig. 4)。这些观察结果强调了秀丽隐杆线虫运动控制的复杂性,以及基于完整结构描述并考虑行为状态和运动输出作为网络涌现属性进行分析的重要性。
5. Reproducibility of connectivity
为了评估连接性的自然变异量,我们检查了雌雄同体重建中,左右同源神经元对神经环中左右同源靶点的连接性。对于化学连接,边缘权重变化了10-40%,具体取决于连接强度(Fig. 5a, Extended Data Fig. 11 and Supplementary Information 8)。缝隙连接的变化更大。个体蠕虫之间的差异预计至少会这么大。以下部分将使用此信息来识别性别差异。
我们检查了一个明显的值得关注的异常值。在两种性别中,味觉神经元ASEL(即该对中的左侧神经元)与嗅觉神经元类别AWC的化学连接性都比ASER(即该对中的右侧神经元)强。已知ASEL-ASER对在感知化学感觉线索的能力上具有侧向性。我们通过对该突触连接进行体内荧光标记,证实了连接性的差异(Fig. 5b)。
6. Comparison of the sexes
秀丽隐杆线虫两种性别的形态和组织的广泛性别分化延伸到了神经系统和末端器官。总共有8个神经元和16块性肌肉是雌雄同体特有的;91个神经元和39块性肌肉是雄性特有的。这些性别特有成分与294个神经元的共享神经系统进行突触整合。在雄性中,67%的共享神经元类别(93个类别中的62个)接收来自性别特有神经元的输入,占其总输入的16%。反过来,54%的共享神经元类别(93个类别中的50个)对性别特有成分有输出,构成性别特有神经元输入的18%。在雌雄同体中,37%的神经元类别被两个性别特有神经元类别HSN和VC靶向,但这仅占共享成分总输入的1.4%。
控制产卵和交配等性别特有行为的回路既包括性别特有的神经元,也包括性别共享的神经元。这在雄性中最为显著,其中控制交配的神经网络由85个雄性特有神经元和64个共享神经元组成。一些共享神经元保留了它们在雌雄同体中相同的功能,但被雄性特有的神经元靶向。例如,主要的1层运动前中间神经元AVA和AVB分别促进后退和前进运动,它们被三个主要的雄性特有中间神经元PVV、PVX和PVY靶向,这些神经元调节雄性在交配过程中的运动。其他参与雄性回路的共享神经元在成年雄性中与成年雌雄同体和幼虫阶段相比,发生了广泛的分化,并在两种性别中发挥不同的功能(Fig. 6a–f)。
性别特有的神经元连接到头部的性别共享中央回路中,具有两个功能:调节明显的生殖活动期间的行为,以及介导性别特异性的、求食性的决策。它们的一些目标,其中几个在两种性别中都被靶向,也接收来自性别共享感觉回路的输入,因此代表了性和非性通路整合的点(Fig. 6g, h)。
一些共享神经元,包括感觉神经元,在性别上发生分化,以支持性别特异性的导航选择。已经描述了11类共享神经元的神经递质表达的性别特异性模式。这种神经元特性的差异延伸到了整个神经系统的连接性。Supplementary Information 9中提供了差异的统计排名。我们通过体内突触标记测试了六个化学连接,并证实每个连接都表现出两种性别之间的差异(Fig. 5c, d and Extended Data Fig. 12)。另外六个连接已由其他人通过实验验证。在性分化过程中发生重塑的神经元的连接中发现的另外33个化学和缝隙连接差异也可能是真实的性别差异(Supplementary Information 9)。在中央通路中,存在涉及似乎在性通路中起作用的共享神经元的性别差异(例如,AVF到RIF和RIM的雄性特异性连接,以及AIM到AIB的雄性特异性连接(AIM显示神经递质的雄性特异性表达,并表达雄性交配搜索促进神经肽PDF-1))。通过对性成熟前幼年动物的荧光突触报告基因的分析,我们进一步发现,除了先前描述的性别特异性修剪外,性别二态性突触也可以通过突触接触的性别特异性生长产生(Fig. 5d)。
仅从电子显微镜重建中识别性别差异是困难的。如上文评估的那样(Fig. 5a),差异可能归因于发育变异、年龄差异、饲养条件或经验,更不用说重建本身中的误差和完整程度的差异。考虑到这些注意事项,差异的统计排名(Supplementary Information 9)提供了一个可供进一步检查的候选列表。即使接受这些保留意见,从上述结果的推断表明,在任何一种重建中发现的 10-30% 的实质性连接(超过三个连续的电镜切片)可能代表了两种性别之间强度差异很大的连接。这种可能的差异涉及化学和缝隙连接,并且发生在整个网络的所有层面上(Extended Data Fig. 13)。
7. Discussion
我们展示了两种成年性别线虫的整个神经系统的物理连接矩阵。这些定量的、相互连接的网络为解析秀丽隐杆线虫行为的神经控制提供了起点。感觉输入通路的汇聚和发散程度使得一般情况下难以轻易识别特定的行为反应通路。主要的运动神经元以及它们的主要运动前中间神经元高度互连,并接受来自剩余大部分神经元的一些输入,这使得对运动输出的简单解释变得困难。此外,结构连接组仅描述了功能通信网络的一部分。通过神经递质、神经肽和激素进行的突触外通信提供了第二个维度,用于控制信息流以实现最佳输出。这里概述的复杂电路必定是秀丽隐杆线虫中许多已知行为的基础,也是那些了解较少或新颖行为(如学习和记忆、动物间交流、社会行为和交配的复杂性)的基础。
这些全动物秀丽隐杆线虫连接组应该被认为是一种概念化,因为它们是由多个动物构建而成,并且包括一些在另一性别中观察到的连接,以及一些根本没有观察到但基于某些区域的重复性而假设存在的连接。由于自然发育的变异,个体在细节上会有所不同(Fig. 5a)。额外的差异将进一步增加这种基本水平的变化。秀丽隐杆线虫的神经突生长和突触的建立发生在整个幼虫发育过程中,并持续到 L4 幼虫阶段和成虫期,并且随着神经系统老化,异位神经突萌发还会继续。电路活动以及因此的经验对连接性的加强已经被证明,但迄今为止研究甚少。个体蠕虫行为差异在多大程度上是由于连接性差异造成的,仍有待探索。连接组过于复杂,不应不受动态、稳态机制的控制;基于电镜的连接组应被视为一个动态结构的快照。
这些结果在两个方面对建模研究很重要。首先,单个神经元更详细的结构,以及单个突触的位置和结构,可能对于理解神经元在回路中的功能至关重要。尽管秀丽隐杆线虫的神经元在很大程度上是等势的,但控制活动的这些神经元的特性,如钙水平,可能是分隔的,正如先前在一个例子中所证明的那样。因此,信号可能不是在所有突触处都是均匀的或同步的。
其次,如果大量的节点或边(例如,代表电耦合的边)缺失,那么就无法认真地开展在连接网络抽象层面上的神经系统功能建模。纳入末端器官至关重要,因为至少在秀丽隐杆线虫的情况下,它们是电耦合的,这将对其活动产生显著影响。连接细胞链中的任何缺口,例如驱动秀丽隐杆线虫运动的运动神经元和肌肉链,都会阻止信号的传播。这里试图描述的连接组足以用于计算分析。
我们对侧下方运动神经元的描述、对中央性通路中中间神经元的识别、对一类在原本分层结构中进行连接的中间神经元的记录,以及对神经系统与非肌肉末端器官(如皮下组织)之间重要界面的描述,都突显了连接组方法对于理解神经系统的功能和行为的重要性。从布罗卡时代开始,一个多世纪以来,神经科学家们一直致力于寻找行为的神经基础。对于这项工作,连接组重建是一种强大的方法。一些神经元的功能可以从它们的连接性中推断出来,这可以将它们识别为以前未知的细胞类别。神经元如何发出信号取决于它们是化学耦合还是电耦合。电镜水平的超微结构可以指示神经元的功能,如感觉感知或激素分泌。建模方法,其完整性至关重要,可以通过连接组来实现,并提供新的见解。物理结构提供了大量的新信息,其中大部分是无法预见的;未知的未知事物可能会显现出来。由结构揭示的新信息为后续通过实验方法进行假设检验提供了基础。
三 论文阅读分析

1. 感觉神经元
有哪些感觉信息输入?
- 特殊化学物质感知:chemo sensory(比如某些致命化学物)
- 气味感知:odor sensory(比如氧气,二氧化碳,社交信号)
- 温度感知:thermo sensory
- 触觉感知:touch sensory
- 痛觉感知:nociception sensory
- 食物感知:food sensory
怎么分类感觉神经元?SN = Sensory Neuron
- SN1:food, nose touch
- SN2:chemo
- SN3:touch
- SN4:O2, CO2, social
- SN5:nociception
- SN6:chemo, odor, thermo
2. 中间神经元
怎么分类中间神经元?IN = Interneuron
- IN1:主要面向运动神经元输出
- IN2:主要面向IN1神经元输出
- IN3:主要面向IN2神经元输出
- IN4:神经元输出没有明显偏向
中间神经元有何特点?
- IN3主要向IN2输出,IN2主要向IN1输出,IN1主要向运动神经元输出
- IN1, IN2, IN3可以接受感觉神经元的输出,可以向运动神经元输出
- IN1, IN2, IN3, IN4相互均存在双向连接,IN4很少向运动神经元输出
- 神经元环路非常普遍,循环反馈调节模式是中间神经元的通路连接特色
- 神经系统的复杂逻辑交互与动态信息处理的核心在于中间神经元连接
3. 运动神经元
有哪些运动信息输出?MU = Muscle
- 头部肌肉:HMU
- 躯体肌肉:BMU(除头部外的其余部分)
怎么分类运动神经元?MN = Motor Neuron
- HMN:头部运动神经元
- BMN:躯体运动神经元
- SMN:姿态运动神经元(控制身体运动姿态)
- VC:ventral nerve cord,腹侧性相关运动神经元
- HSN:hermaphrodite-specific neurons,雌雄同体性相关运动神经元
雌雄同体和雄性线虫的神经元系统主要不同在于性相关神经元的差异,性相关神经元主要两类:VC和HSN。雌雄同体的精子进入和产卵通道在腹侧中部,雄性精子输出主要在尾部。
4. 全部神经元

简化图:

雌雄同体:

雄性个体:

5. 相关的数据
神经元分类-秀丽隐杆线虫
神经元连接-雌雄同体线虫
神经元连接-雄性个体线虫
未来的人工智能注定分化为两条路:一条路以连续计算为主体的拟合智能,以深度学习为代表;另一条路以离散计算为主体的形式智能,以生物智能为代表。客观说,并没有任何一种算法可以解决所有类型问题,每种算法都有其适用的领域。
连续计算为主的智能优点在于建模简单,缺点在于无法高效分离计算导致复杂度较高;离散计算为主的智能优点在于可以有效分离计算从而降低复杂度,缺点在于难以有效建立学习模型。两者在实现智能上均存在各自的优势,也许相互协作才是最终的归宿。
相关文章:
论文阅读:《Whole-animal connectomes of both Caenorhabditis elegans sexes》
一 论文整体概述 论文下载链接:《Whole-animal connectomes of both Caenorhabditis elegans sexes》 补充信息和额外数据:https://www.nature.com/articles/s41586-019-1352-7 1. 作者期刊背景 该论文由Scott W. Emmons,David H. Hall等…...

嵌入式开发之STM32学习笔记day03
STM32之ADC(模拟数字转换器) 1 ADC简述2 ADC转换时间3 ADC转化结果存放机制4 ADC转化结果存放机制5 ADC电压转换 1 ADC简述 ADC(Analog-Digital Converter)模拟—数字转换器;ADC可以将引脚上连续变化的模拟电压转换为…...

windows10 安装 Golang 版本控制工具g与使用
下载包:https://github.com/voidint/g/releases 解压, 并添加到环境变量 g 常用命令 查询当前可供安装的stable状态及所有的 go 版本 # stable 版本 g ls-remote stable# 所有版本 g ls-remote安装目标 go 版本1.23.4g install 1.23.4切换到已安装的…...
SpringBoot 使用 Cache 集成 Redis做缓存保姆教程
1. 项目背景 Spring Cache是Spring框架提供的一个缓存抽象层,它简化了缓存的使用和管理。Spring Cache默认使用服务器内存,并无法控制缓存时长,查找缓存中的数据比较麻烦。 因此Spring Cache支持将缓存数据集成到各种缓存中间件中。本文已常…...

R数据分析:多分类问题预测模型的ROC做法及解释
有同学做了个多分类的预测模型,结局有三个类别,做的模型包括多分类逻辑回归、随机森林和决策树,多分类逻辑回归是用ROC曲线并报告AUC作为模型评估的,后面两种模型报告了混淆矩阵,审稿人就提出要统一模型评估指标。那么肯定是统一成ROC了,刚好借这个机会给大家讲讲ROC在多…...

数据结构与算法之二叉树: LeetCode 654. 最大二叉树 (Ts版)
最大二叉树 https://leetcode.cn/problems/maximum-binary-tree/ 描述 给定一个不重复的整数数组 nums 。 最大二叉树 可以用下面的算法从 nums 递归地构建: 创建一个根节点,其值为 nums 中的最大值递归地在最大值 左边 的 子数组前缀上 构建左子树递归地在最大值…...

Linux 容器漏洞
定义:Linux 容器漏洞是指在容器技术(如 Docker、LXC 等)运行环境中存在的安全弱点。这些漏洞可能存在于容器镜像本身、容器运行时(如 runc)、容器编排工具(如 Kubernetes)或者容器与主机之间的交…...
)
file与io流(1)
-1- java.io.File类的使用 (1) 概述 File类及本章下的各种流,都定义在java.io包下。一个File对象代表硬盘或网络中可能存在的一个文件或者文件目录(俗称文件夹),与平台无关。(体会万事万物皆…...

忘记了PDF文件的密码,怎么办?
PDF文件可以加密,大家都不陌生,并且大家应该也都知道PDF文件有两种密码,一个打开密码、一个限制编辑密码,因为PDF文件设置了密码,那么打开、编辑PDF文件就会受到限制。忘记了PDF密码该如何解密? PDF和offi…...

Linux权限管理(用户和权限之间的关系)
Linux系列 文章目录 Linux系列一、Linux下用户类型二、普通权限的基本概念2.1、Linux中权限的类别2.2、Linux中权限对应的三种身份2.3、文件权限的标识 三、文件权限设置四、修改文件属主和属组4.1、chown修改文件的属主4.2、修改所属组 五、文件掩码六、目录权限 一、Linux下用…...
Python Selenium库入门使用,图文详细。附网页爬虫、web自动化操作等实战操作。
文章目录 前言1 创建conda环境安装Selenium库2 浏览器驱动下载(以Chrome和Edge为例)3 基础使用(以Chrome为例演示)3.1 与浏览器相关的操作3.1.1 打开/关闭浏览器3.1.2 访问指定域名的网页3.1.3 控制浏览器的窗口大小3.1.4 前进/后…...
【Uniapp-Vue3】使用defineExpose暴露子组件的属性及方法
如果我们想要让父组件访问到子组件中的变量和方法,就需要使用defineExpose暴露: defineExpose({ 变量 }) 子组件配置 父组件配置 父组件要通过onMounted获取到子组件的DOM 传递多个属性和方法 子组件 父组件...
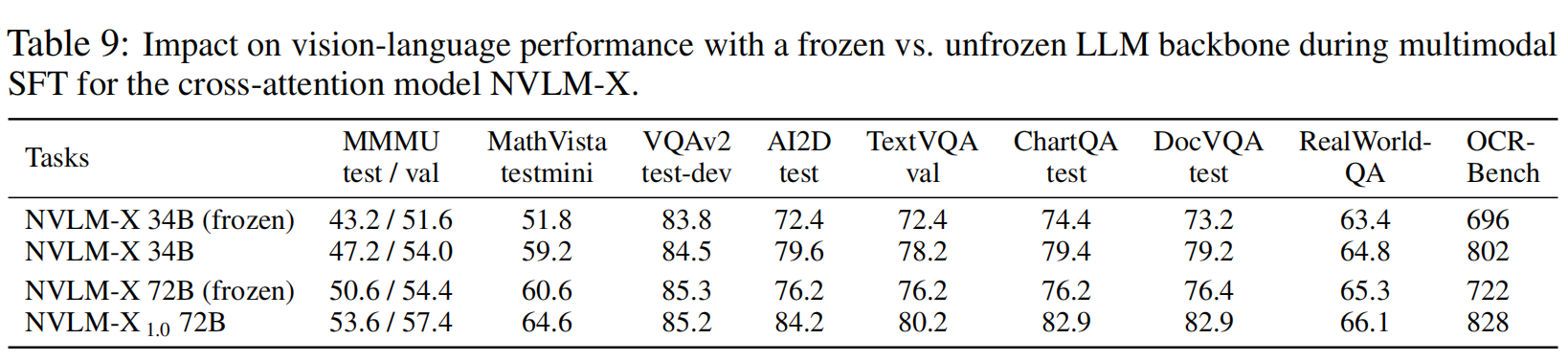
【多模态LLM】英伟达NVLM多模态大模型训练细节和数据集
前期笔者介绍了OCR-free的多模态大模型,可以参考:【多模态&文档智能】OCR-free感知多模态大模型技术链路及训练数据细节,其更偏向于训练模型对于密集文本的感知能力。本文看一看英伟达出品的多模态大模型NVLM-1.0系列,虽然暂未…...

HTTP详解——HTTP基础
HTTP 基本概念 HTTP 是超文本传输协议 (HyperText Transfer Protocol) 超文本传输协议(HyperText Transfer Protocol) HTTP 是一个在计算机世界里专门在 两点 之间 传输 文字、图片、音视频等 超文本 数据的 约定和规范 1. 协议 约定和规范 2. 传输 两点之间传输…...

MySQL教程之:输入查询
如上一节所述,确保您已连接到服务器。这样做本身不会选择任何要使用的数据库,但没关系。在这一点上,了解一下如何发出查询比直接创建表、加载数据和从中检索数据更重要。本节介绍输入查询的基本原则,使用几个查询,您可…...
docker+ffmpeg+nginx+rtmp 拉取摄像机视频
1、构造程序容器镜像 app.py import subprocess import json import time import multiprocessing import socketdef check_rtmp_server(host, port, timeout5):try:with socket.create_connection((host, port), timeout):print(f"RTMP server at {host}:{port} is avai…...
不同音频振幅dBFS计算方法
1. 振幅的基本概念 振幅是描述音频信号强度的一个重要参数。它通常表示为信号的幅度值,幅度越大,声音听起来就越响。为了更好地理解和处理音频信号,通常会将振幅转换为分贝(dB)单位。分贝是一个对数单位,能…...
【17. 电话号码的字母组合 中等】
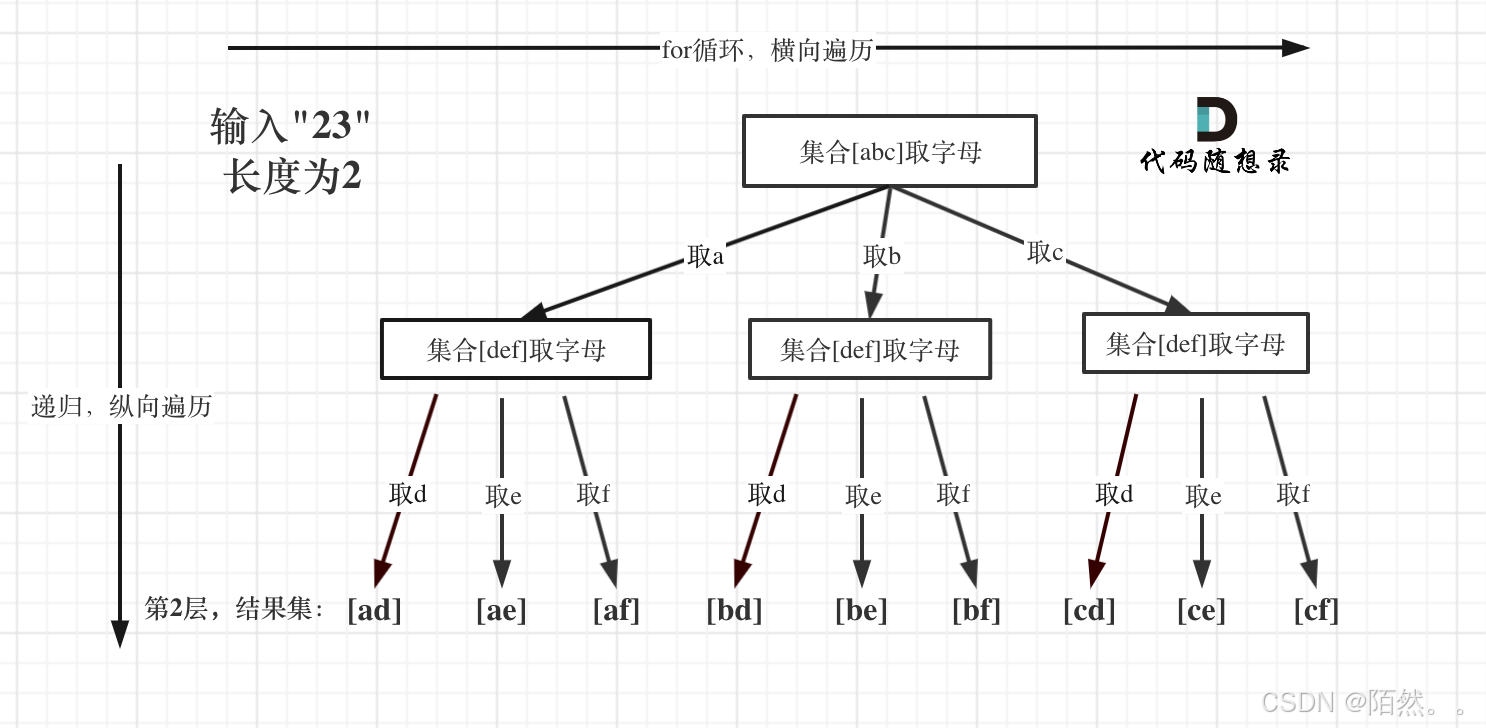
题目: 给定一个仅包含数字 2-9 的字符串,返回所有它能表示的字母组合。答案可以按 任意顺序 返回。 给出数字到字母的映射如下(与电话按键相同)。注意 1 不对应任何字母。 示例 1: 输入:digits “23”…...
数据结构初阶---排序
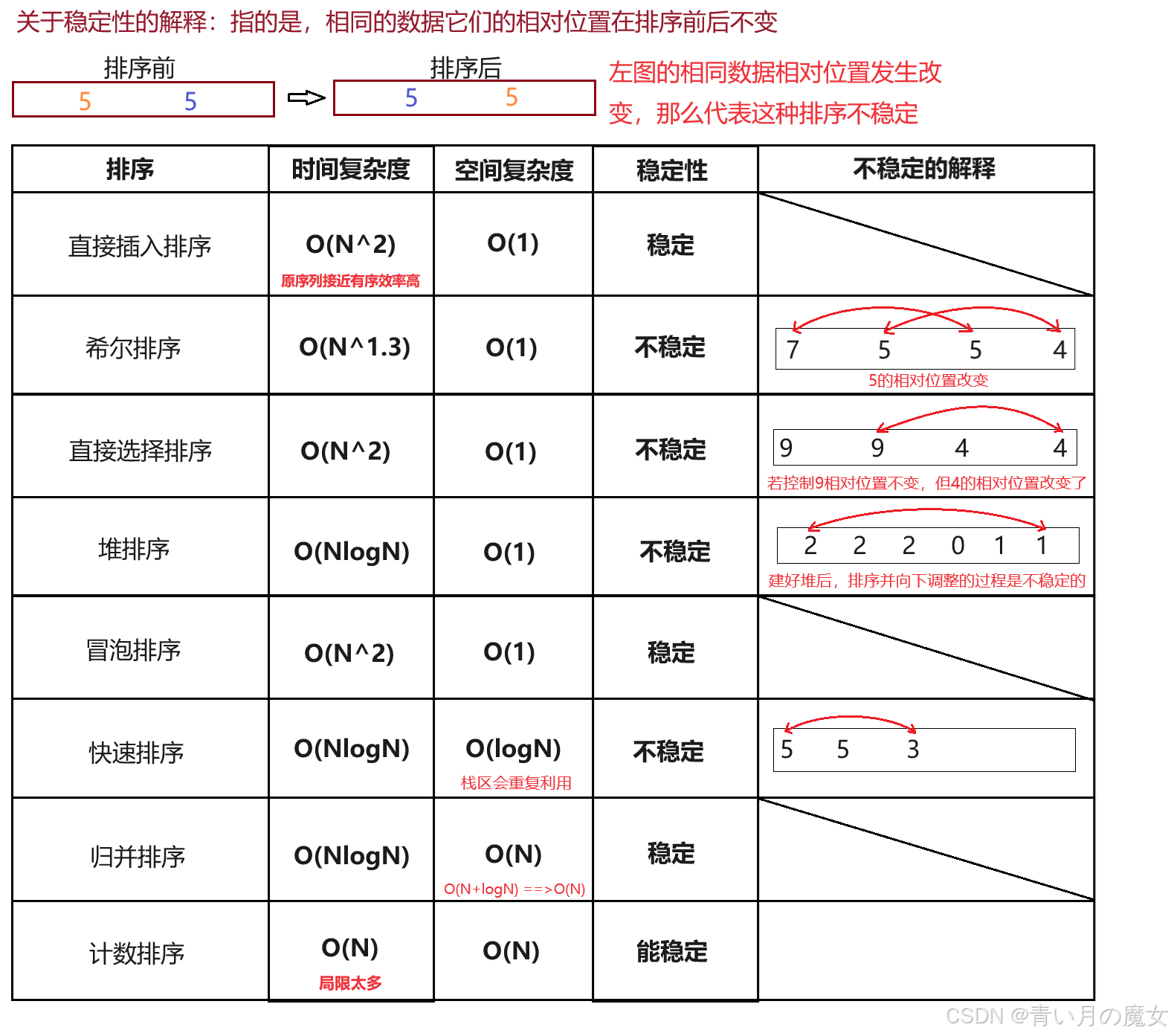
一、排序相关概念与运用 1.排序相关概念 排序:所谓排序,就是使一串记录,按照其中的某个或某些关键字的大小,递增或递减的排列起来的操作。 稳定性:假定在待排序的记录序列中,存在多个具有相同的关键字的…...
【从0-1实现一个前端脚手架】
目录 介绍为什么需要脚手架?一个脚手架应该具备哪些功能? 脚手架实现初始化项目相关依赖实现脚手架 发布 介绍 为什么需要脚手架? 脚手架本质就是一个工具,作用是能够让使用者专注于写代码,它可以让我们只用一个命令…...

DIY复刻经典:Texar Audio Prism动态处理器克隆套件全攻略
1. 项目概述:Texar Audio Prism 克隆套件如果你在专业音频圈子里混过一段时间,尤其是对上世纪八九十年代那些经典的、带点“魔法”色彩的外置动态处理器感兴趣,那么“Texar Audio Prism”这个名字你大概率不会陌生。它不是最常见的1176或者LA…...

[智能体-69]:重新认知MCP:协议不生产智能,只是AI全域交互的标准化基石
MCP只是提供了大模型、编排调度、外部工具能够进行结构化交流的标准,而整个系统的智能主要依赖编排调度,与外部软件系统的交互取决于外部工具,包括外部语音交互、视觉交互、数字化交互。当下MCP(Model Context Protocol࿰…...

ARMv8 HFGITR_EL2寄存器解析与虚拟化指令陷阱控制
1. AArch64 HFGITR_EL2寄存器架构解析HFGITR_EL2(Hypervisor Fine-Grained Instruction Trap Register)是ARMv8架构中专门用于指令级陷阱控制的系统寄存器,属于虚拟化扩展的重要组成部分。这个64位寄存器通过位映射机制实现对特定AArch64指令…...

一次搞懂内存取证:用Volatility3和Cobalt Strike分析工具复现VNCTF‘来一把紧张刺激的CS’
实战内存取证:从Volatility3到Cobalt Strike信标分析全解析 在网络安全事件响应中,内存取证往往是发现高级威胁的最后一道防线。当攻击者使用文件无落地的技术时,传统的磁盘取证可能一无所获,而内存中却保留着攻击行为的完整痕迹。…...

第三卷第4章:原型模式设计思想
第三卷第4章:原型模式设计思想 目录介绍 01.案例引入与思考 1.1 痛点场景 1.2 它哪里不舒服 1.3 引出本篇主角 02.原型模式介绍 2.1 原型模式由来 2.2 原型模式定义...

Python UiAutomation实战:从网页数据抓取到桌面应用,一个库打通数据采集全链路
Python UiAutomation实战:打通数据采集全链路的智能解决方案 在数据驱动的商业环境中,企业常常面临跨平台数据采集的挑战——财务系统里的交易记录需要与网站后台的报表进行交叉分析,销售数据要从桌面软件导出后上传到云端处理系统。传统的人…...

Godot 2D随机地图三大静默故障:黑屏、穿墙、寻路失败的根源与修复
1. 为什么刚上手Godot做2D随机地图就总卡在“生成出来是黑的”“角色穿墙”“房间连不通”这三件事上?如果你是刚从Unity或GameMaker转来Godot,或者第一次用GDScript写程序逻辑的新手,大概率已经在2D随机地图生成这个环节反复摔过跟头——不是…...

智能体任务分配算法:从启发式到深度强化学习的演进与实践
1. 项目概述:从“谁来做”到“如何做得更好”的智能进化在机器人集群、无人机编队或是自动化仓储系统中,我们常常面临一个看似简单实则复杂的问题:眼前有一堆任务,手头有一群可用的智能体(机器人、无人机、服务器等&am…...

终极Obsidian笔记模板指南:如何用kepano-obsidian构建你的第二大脑
终极Obsidian笔记模板指南:如何用kepano-obsidian构建你的第二大脑 【免费下载链接】kepano-obsidian My personal Obsidian vault template. A bottom-up approach to note-taking and organizing things I am interested in. 项目地址: https://gitcode.com/gh_…...
)
H3C VSR路由器实战:用QoS策略给不同VLAN用户打DSCP标签(附配置命令详解)
H3C VSR路由器QoS实战:基于VLAN的DSCP标记与流量调度指南 在企业网络环境中,不同业务对网络质量的需求差异显著。普通办公流量可以容忍轻微延迟,但视频会议需要稳定的低延迟保障,而访客上网则可能消耗大量带宽却无需优先保障。本文…...